Key Result
CRISPR technology was used to generate a range of starch branching enzyme (SBE) mutants in canola. Sbe quadruple and sextuple mutants were used for expressing maize endosperm SBEI and effects on growth, morphology, flowering and yield determined. Introduction of corn SBEI into quadruple mutants (four canola genes edited) repeatedly led to increased total seed yield of 30 per cent in greenhouse conditions. In the sextuple sbe mutant an additional phenotype of a thicker stem conferred improved resistance to drought and high temperature. Additional lines are being generated to test under field conditions and a patent has been granted.
Project Summary
Introduction
Carbohydrates such as starch provide the stored energy reserves of plants. The goal of the plant is to ensure seed production and the research team previously developed a novel technology which caused a remarkable boost in seed yield in Arabidopsis by modifying starch metabolism. When the Arabidopsis endogenous leaf starch branching enzymes (SBEs) were replaced with maize endosperm homologues ZmSBEI or ZmSBEIIb, the Arabidopsis plants demonstrated significant increases in starch biosynthesis and a dramatic increase in seed production that led to a 250% increase in total seed oil produced per plant (Liu et al., 2016). The increase in seed production was associated with an increase in the numbers of flowers and siliques per plant, while the fatty acid profile of the seed oil remained unaffected.
Canola (Brassica napus) is genetically close to Arabidopsis with highly conserved gene functions between the two species. The homologous SBEs in canola are assembled on both A and C genomes and they have high identities to those in Arabidopsis. This provided a feasible strategy to apply the above technology to canola. Canola is allotetraploid with a more complicated genetic background and, since no SBE knockout mutants are so far publicly available, the strategy for replication of this effect in canola has been divided into two stages:
(1) deletion of endogenous BnaSBEs by gene editing;
(2) expression of maize SBEI or SBEIIb.
Gene editing using the CRISPR/Cas9 system has been applied to edit the endogenous SBEs and the researchers successfully produced homozygous mutant lines targeting six SBE genes. The sbe sextuple mutant was analysed with respect to biochemistry and plant development. Characterization of positive lines expressing maize SBEI or SBEIIb in the sbe sextuple mutant background was accomplished.
Milestones and deliverables achieved:
(1) evaluated the T1 generation of canola lines expressing ZmSBEI in the sextuple sbe mutant; these lines show an 8-20% increase in total seed yield, which is very promising given that they are a mix of heterozygotes and homozygotes;
(2) identification of homozygous lines for further evaluation and field trials;
(3) characterization of the “thick” main stem phenotype in the sbe sextuple mutant when compared to wild type plant;
Details of (1-3) are presented in the following sections.
Results:
(1) Effects of expression of ZmSBEI in the sbe sextuple mutant
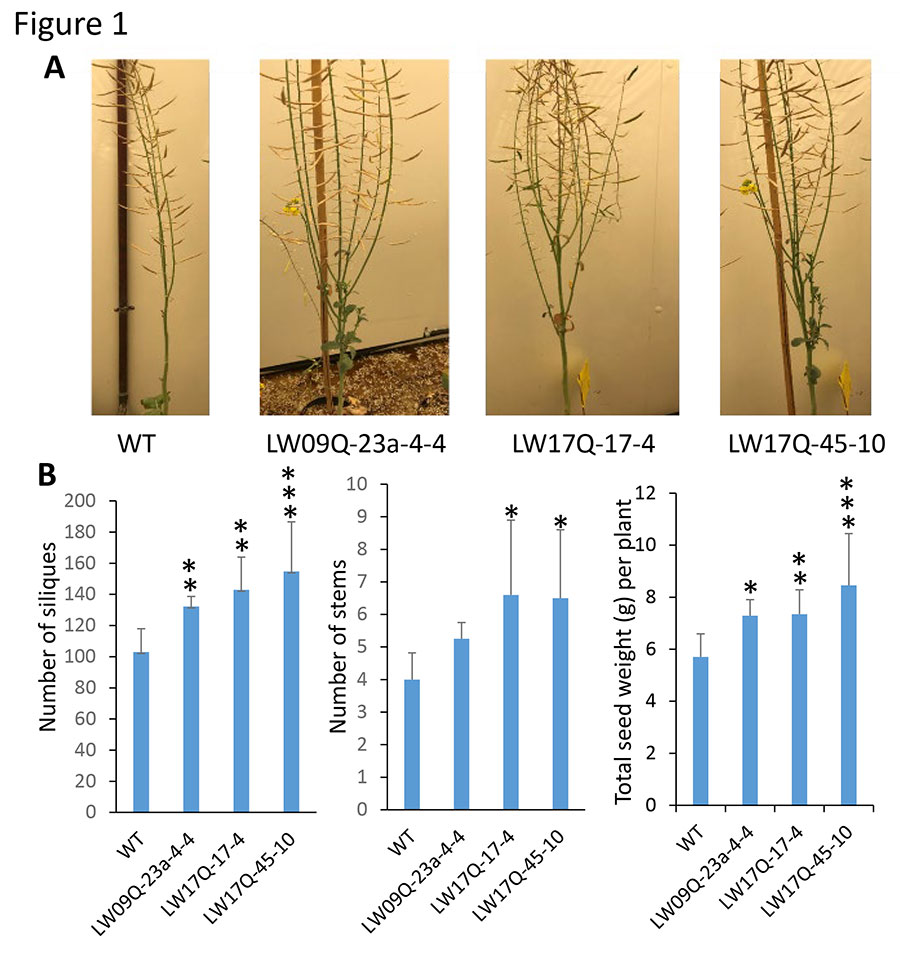
Having shown that the number of flowers in the lines expressing ZmSBEI in the sbe sextuple mutant was increased up to 2-fold compared to the WT, the research team then analyzed the number of siliques and total seed weight. Even at this early stage, with many more transgenic plants in the pipeline, line LW17S-1 displayed increased silique number (10-30%) and increased total seed weight (8-20%) (Figure 1). Three more independent lines are at the early stage of T1 generation and await phenotype evaluation. These data are highly promising and consistent with the results of ZmSBEI expression in the sbe quadruple mutant background.
(2) Identification of homozygous lines expressing ZmSBEI in the sbe sextuple mutant
Since the previously selected T0 lines expressing ZmSBEI in the sbe sextuple mutant were shown to contain a single T-DNA insertion, those T1 plants are expected to be homozygous when their progeny are all positive and do not segregate. Thus, homozygous lines have now been identified from three, independent, T0 lines, and their phenotypes will be further evaluated in greenhouse and field.
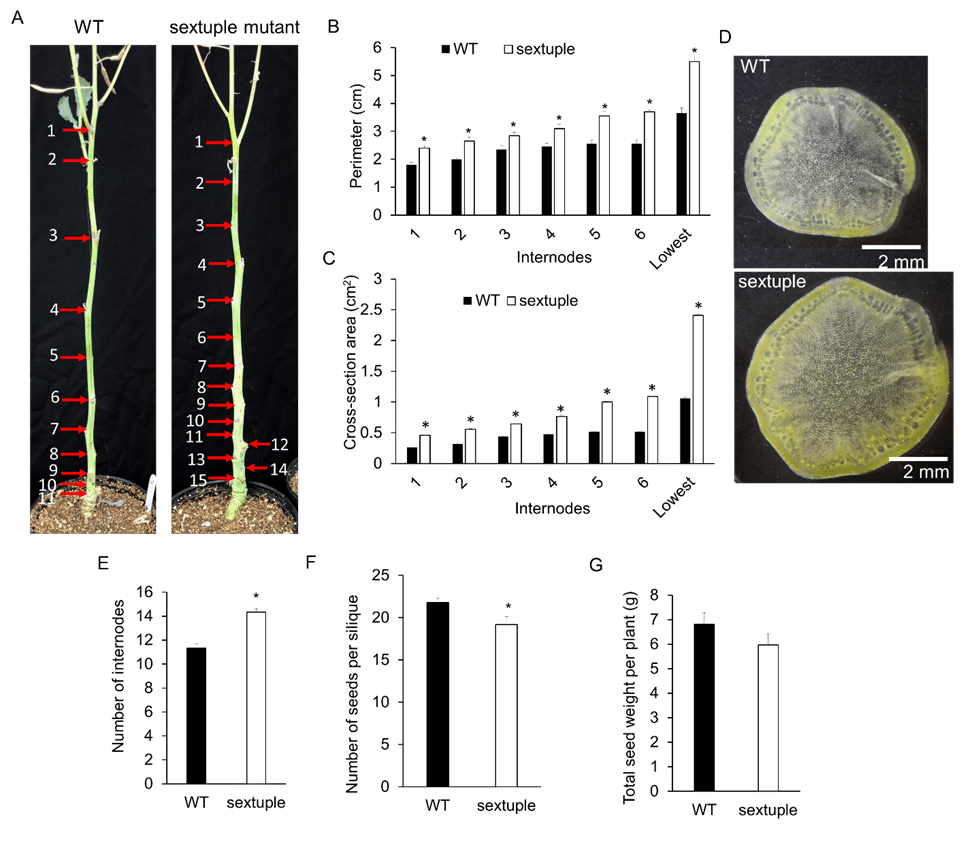
(3) Characterization of the thick main stem phenotype in the sbe sextuple mutant While phenotyping the sbe sextuple mutant and WT plants, the research team noticed that at the maturation stage, the main stem in the sextuple mutant was evidently thicker than WT, by 21-50% when measuring the perimeter of internodes (Figure 2A, B). The cross-section areas in each internode was calculated using their respective perimeters, showing remarkably increased areas in the sextuple mutant by 80-125% (Figure 2C). For illustration, cross-sections of the second internode are shown in Figure 2D. There is clearly an increase in stem diameter and vascular bundle area in the sextuple mutant compared to WT. The research team also found that the sextuple mutant developed on average three more internodes than WT plants (Figure 2A, E). The additional sextuple mutant, LW13Q-22-167-8, which contains different mutations in BnaA10_SBE2.1 genes from the current line LW13Q-22-59-36, also displayed this phenotype consistent with the hypothesis that the thick stem is associated with BnaSBE mutations. The seed number per silique in the sextuple mutant was approximately 10% less than that in WT (Figure 2E), resulting in slightly reduced total seed weight (10%) in the sextuple mutant (Figure 2F). Importantly, initial observations suggest that transgenic lines expressing ZmSBEI in the sextuple mutant produce a similarly thickened stem as well as increased yield compared to WT, suggesting a “double benefit” from these genetic changes.
This research resulted in the publication of CRISPR–Cas9-mediated editing of starch branching enzymes results in altered starch structure in Brassica napus.
Future directions
The research team will continue to analyze the transformants expressing ZmSBEI or ZmSBEIIb in the sbe sextuple mutant background. Previous reports have shown that high temperature at flowering causes a deterioration in stem mechanical properties in canola, resulting in increased risk of crop lodging and accompanied yield loss (Wu et al., 2021, J Agro Crop Sci 207: 74-87). Given that the sbe sextuple mutant seeds maintained wildtype levels of seed oil synthesis (previous report), the research team will test whether the thickened stems can minimize yield loss under high temperature/drought stress in controlled environmental conditions. More importantly, our preliminary data has shown that the lines expressing ZmSBEI/ZmSBEIIb also possess the thick stem phenotype. Thus, once there are a number of homozygous lines expressing ZmSBEI or ZmSBEIIb in the sbe sextuple mutant background, the research team will test whether the higher yield performance of these transgenic lines, compared to WT, is further enhanced during abiotic stress prior to testing in field conditions. Given the anticipated changes arising from global climate change this technology may provide additional advantages. Once the current analyses are completed, the research team will look to secure further funding to undertake these tests in greenhouse and field trials.
The Evaluation of gene-edited canola with improved yield and abiotic stress tolerance project builds on the findings of this research.
References
- Liu F, Zhao Q, Mano N, Ahmed Z, Nitschke F, Cai Y, Chapman KD, Steup M, Tetlow IJ, Emes MJ (2016) Modification of starch metabolism in transgenic Arabidopsis thaliana increases plant biomass and triples oilseed production. Plant Biotechnol J 14: 976–985.
- Wu W, Duncan RW, Ma B-L (2021) The stage sensitivity of short-term heat stress to lodging-resistant traits and yield determination in canola (Brassica napus L.). J Agro Crop Sci 207: 74–87.